

信号肽对蛋白表达量的影响及其在纯化中的作用解析
- 2026-06-23
- 298
重组蛋白的高效表达与低成本纯化是生物医药、工业酶制剂研发生产的核心痛点,信号肽作为新生蛋白N端15-30个氨基酸的短肽序列,是介导蛋白跨膜转运、调控表达效率、简化纯化工艺的关键分子元件。
信号肽的序列结构与宿主转运系统的适配性直接决定重组蛋白的分泌水平、可溶性与生物活性,不仅能有效规避胞内表达易形成无活性包涵体的问题,还可通过定向分泌大幅降低下游纯化的杂质干扰与工艺成本,是串联基因构建、表达调控与纯化制备全流程的核心分子工具。
一、信号肽的结构基础
信号肽的功能核心源于保守的N-H-C三区域结构,该结构是其被转运系统识别、介导跨膜、完成切割的分子基础,不同物种与蛋白的信号肽序列虽有差异,但结构框架高度统一。
N区为信号肽N端碱性区域,由精氨酸、赖氨酸等带正电荷氨基酸组成,净正电荷通常为+2-+3,主要负责与细胞膜磷脂双层、信号识别颗粒(SRP)结合,启动蛋白转运进程。革兰氏阳性菌信号肽N区长度与正电荷显著高于革兰氏阴性菌,真核生物N区则更短小紧凑。
H区是信号肽疏水核心,由7~15个亮氨酸、异亮氨酸等疏水氨基酸构成,可形成稳定α-螺旋构象,是信号肽嵌入细胞膜、选择转运通路的关键区域。其疏水性强弱直接决定蛋白转运效率,过度疏水会导致跨膜阻滞,疏水不足则无法完成膜定位。
C区为信号肽切割区域,含信号肽酶识别的保守位点,遵循**(-3,-1)规则**,经典基序为Ala-X-Ala,长度3~7个氨基酸,负责信号肽精准切割,保障成熟蛋白的正确生成。
此外,信号肽按转运机制可分为Sec/SPI、Sec/SPII、Tat/SPI等5类,其中Sec依赖型信号肽是原核表达系统最常用类型,如金黄色葡萄球菌蛋白A(SpA)信号肽,经改造后可适配大肠杆菌Sec通路,高效介导异源蛋白分泌。
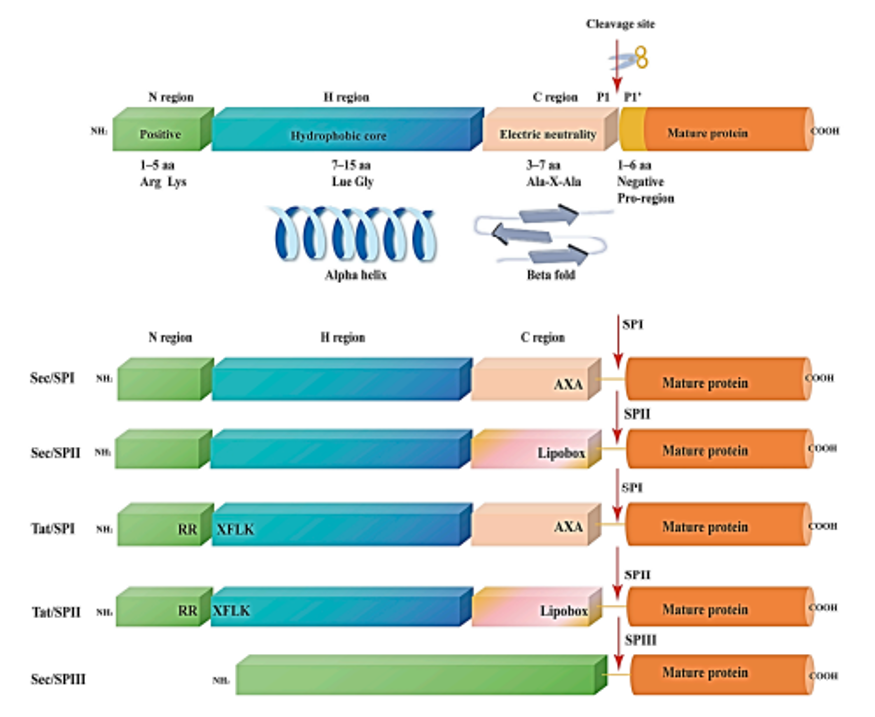
图1 经典N-H-C三区域信号肽结构
二、调控蛋白表达的机制
信号肽通过转运定位、翻译调控、折叠保护三重机制,直接调控重组蛋白的表达量、可溶性与活性,是避免包涵体形成、提升活性蛋白产量的核心要素。
在翻译调控层面,信号肽与宿主密码子偏好性、翻译系统的兼容性,直接决定mRNA稳定性与翻译效率。hGH的天然真核信号肽无法被大肠杆菌Sec/Tat通路识别,会直接导致翻译阻滞、蛋白合成量骤降,胞内大量形成无活性包涵体。
在转运定位层面,信号肽通过SRP、Sec、Tat三条通路介导蛋白转运:SRP通路为共翻译转运,同步完成翻译与跨膜,杜绝蛋白胞内错误折叠;Sec通路为后翻译转运,负责未折叠蛋白跨膜运输;Tat通路则转运已折叠蛋白。信号肽疏水性决定通路偏好,高疏水偏好SRP通路,中等疏水适配Sec通路,弱疏水则通过Tat通路。
在折叠保护层面,信号肽介导的周质/胞外分泌,可利用宿主周质空间的二硫键异构酶、肽酰脯氨酸顺反异构酶辅助蛋白正确折叠,减少胞内蛋白酶降解,大幅提升活性蛋白比例。改造后的SpA信号肽可将hGH定向转运至大肠杆菌周质与胞外,活性蛋白占比达28.12%,远高于胞质表达的10%。
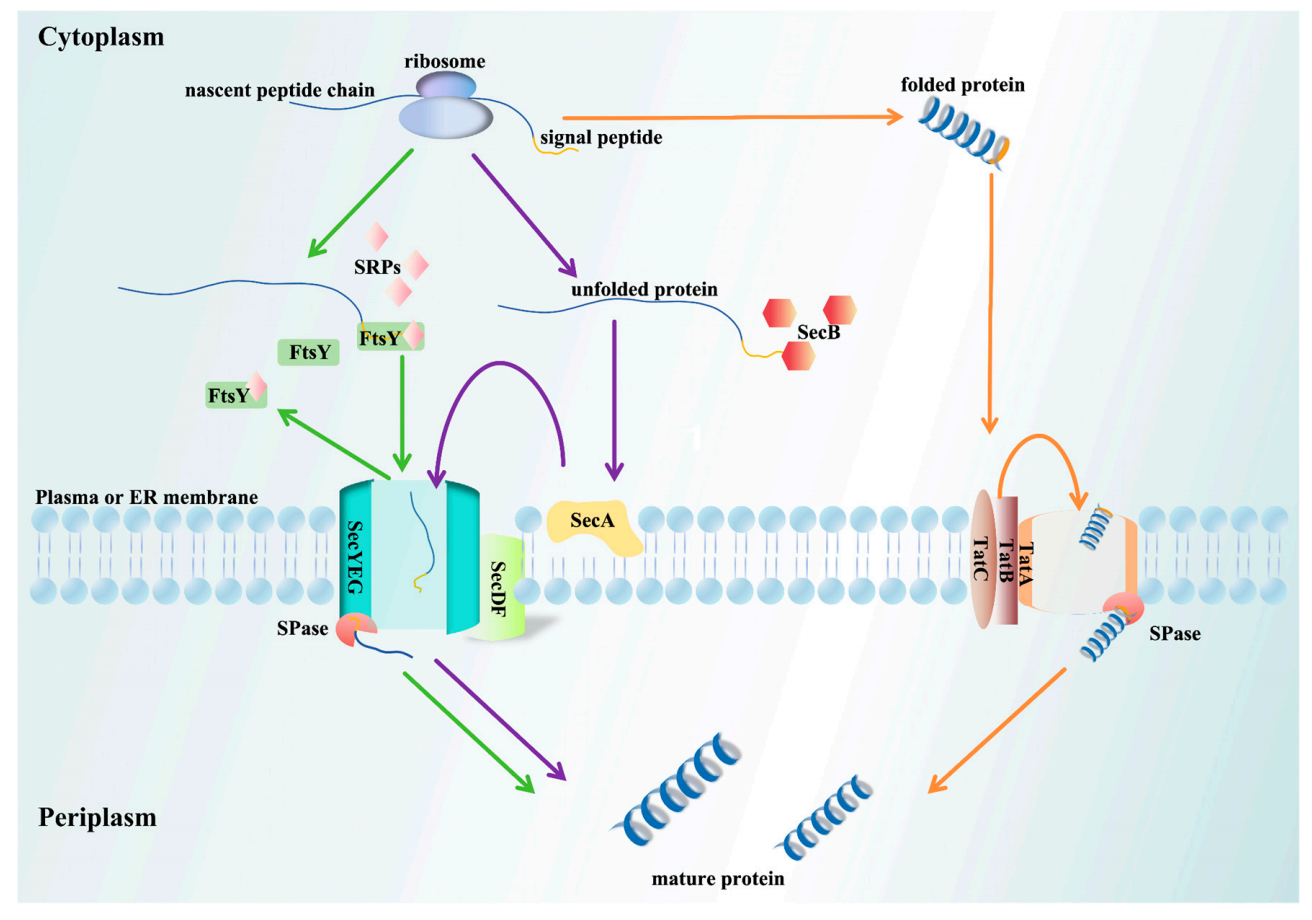
图2 信号肽介导的蛋白跨膜转运通路图
三、提升表达量的优化策略
信号肽的理性改造与配套工艺优化,是突破重组蛋白表达瓶颈的核心手段,结合生物信息学设计与实验验证,可实现表达效率最大化。
(1)结构定向改造。N区优化以适度增加正电荷为核心,将SpA信号肽N区亮氨酸替换为精氨酸,提升与细胞膜静电结合力;H区通过替换亲水性氨基酸为疏水氨基酸(苏氨酸→异亮氨酸、丝氨酸→丙氨酸),强化α-螺旋稳定性,同时避免疏水度过高导致转运阻滞;C区严格遵循Ala-X-Ala保守基序,优化切割位点,杜绝信号肽变体(SPVs)产生。
(2)密码子与框架优化。对信号肽与目的蛋白进行宿主密码子优化,hGH基因经大肠杆菌密码子优化后,密码子适应指数从0.85提升至0.87,GC含量稳定在50.59%,消除mRNA茎环结构,翻译效率显著提升;同时在起始密码子前添加Kozak序列,强化翻译起始效率。
(3)表达工艺优化。采用响应面法(RSM)确定最优诱导条件:IPTG浓度0.6mM、诱导温度26℃、菌体OD600=1.0时诱导、诱导时间10h,可最大化hGH分泌表达量;添加1%-1.5%甘氨酸提升细胞膜通透性,进一步促进蛋白胞外分泌。
(4)宿主适配筛选。构建信号肽文库高通量筛选适配元件,天然SpA信号肽无法介导hGH分泌,经改造后则可实现高效分泌,打破天然信号肽的宿主兼容性限制。
四、简化纯化的核心作用
信号肽介导的分泌型表达,从流程简化、纯度提升、活性保留三个维度,重构重组蛋白纯化工艺,大幅降低工业化生产成本。
一是简化前处理流程。胞内表达需破碎细胞释放蛋白,易引入宿主杂蛋白、内毒素、核酸污染;信号肽介导的胞外/周质分泌,可直接收集培养上清或周质组分,无需细胞破碎,省去裂解、离心除碎片等步骤,纯化流程缩短50%以上。
二是降低污染提升纯度。大肠杆菌胞质内毒素与宿主蛋白含量极高,分泌至胞外的蛋白上清中杂蛋白占比极低,内毒素污染水平显著下降,仅需一步Ni-NTA亲和层析即可获得纯度95%以上的目标蛋白,无需多步层析纯化。
三是保留蛋白天然活性。分泌表达的蛋白无需包涵体复性,避免复性过程中蛋白变性、聚集,且成熟蛋白N端无多余甲硫氨酸,降低免疫原性;hGH周质分泌产物的生物活性与天然蛋白高度一致,远优于胞质表达的复性蛋白。
五、应用展望
信号肽已成为原核、真核重组蛋白表达系统的核心元件,广泛应用于生物医药、疫苗研发等领域,未来应用边界将持续拓展。
在工业生产领域,大肠杆菌OmpA、PelB信号肽,枯草芽孢杆菌AmyE信号肽,已用于hGH、胰岛素等治疗性蛋白的工业化分泌生产;酵母α-因子、人血清白蛋白信号肽适配哺乳动物细胞,大幅提升单克隆抗体的表达与分泌效率。
在疫苗研发领域,信号肽可作为内源性佐剂,提升抗原分泌与MHCI类分子提呈效率,新冠mRNA疫苗采用tPA信号肽,显著提升刺突蛋白表达量与免疫原性;DNA疫苗、病毒载体疫苗通过融合信号肽,可增强CD8+T细胞应答,提升疫苗保护效力。
未来发展中,机器学习工具(SignalP6.0)将实现信号肽功能的精准预测,高通量筛选与合成生物学结合,可设计通用型高效信号肽;信号肽与环状RNA(circRNA)疫苗的融合,能延长抗原表达时间,进一步提升疫苗稳定性与保护效果。此外,信号肽在免疫调控、遗传疾病筛查中的非经典功能挖掘,将为生物医药研发提供新方向。
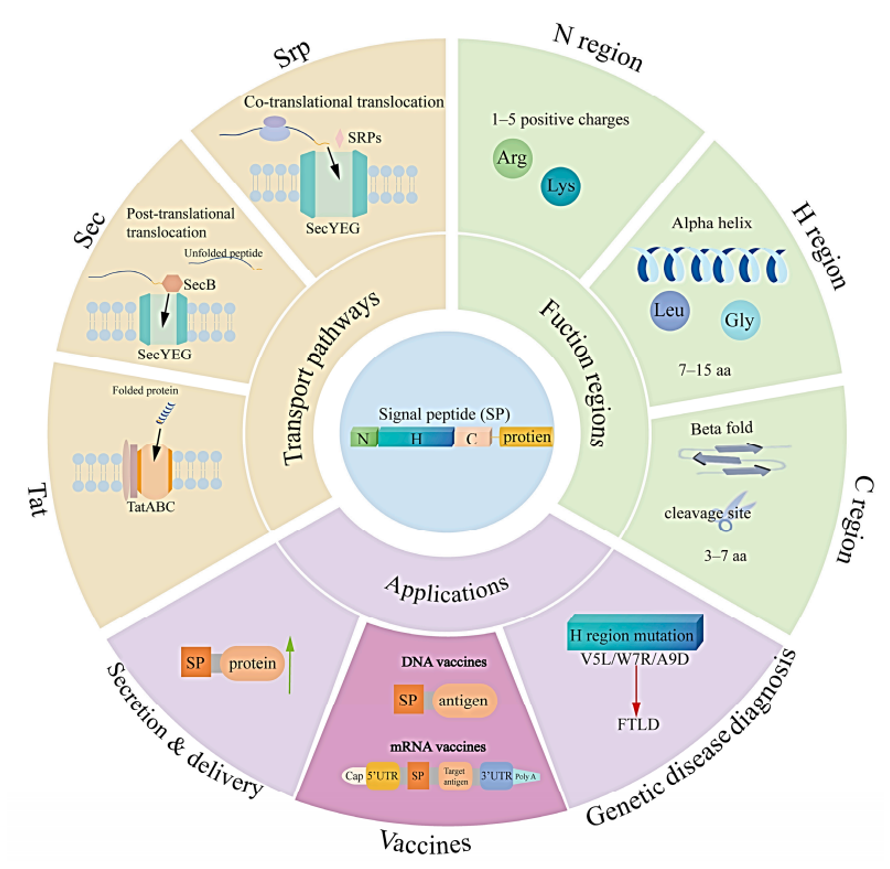
图3 信号肽的结构、转运通路及应用场景
卡梅德生物(KMD Bioscience)依托成熟的蛋白研发与制备平台,可完成小分子、多肽、蛋白等多样化抗原的设计与修饰,确保高免疫原性。基于杂交瘤与噬菌体展示技术,可提供鼠、兔、羊驼等多物种的单克隆抗体制备服务。同时,高活性蛋白可用于肽库筛选,包括固相、液相及细胞筛选等策略,通过高效淘选快速获得高亲和力序列,为抗体药物研发、诊断试剂开发及基础研究提供专业可靠的技术支撑。
[1] Zhang S, He Z, Wang H, Zhai J. Signal Peptides: From Molecular Mechanisms to Applications in Protein and Vaccine Engineering. Biomolecules. 2025;15(6):897.
[2] Rigi G, Rostami A, Ghomi H, et al. Optimization of expression, purification and secretion of functional recombinant human growth hormone in Escherichia coli using modified staphylococcal protein a signal peptide. BMC Biotechnol. 2021;21(1):51.
[3] Duan G, Ding L, Wei D, et al. Screening endogenous signal peptides and protein folding factors to promote the secretory expression of heterologous proteins in Pichia pastoris. J Biotechnol. 2常见问题汇总
Q1:什么是信号肽?它如何从机制上影响蛋白表达量?
A:信号肽是新生蛋白N端一段长度为15–30个氨基酸的短肽,通常由带正电的N区、疏水的H区和极性的C区构成。它通过引导核糖体结合至内质网膜上的Sec易位子,将翻译中的多肽链送入内质网腔,从而启动分泌途径。
信号肽对表达量的影响主要体现在三个方面:
第一,它促进蛋白向细胞外或周质空间转运,避免胞内错误折叠或形成包涵体,从而显著提高可溶性表达水平。例如,在大肠杆菌中,使用PelB信号肽可将胞质中不溶的蛋白转变为周质空间的可溶形式,表达量提升数倍。
第二,信号肽介导的共翻译转运能减缓翻译延伸速率,为新生肽链提供更充裕的折叠时间,降低聚集倾向。
第三,不同宿主细胞对信号肽的识别效率差异巨大。在真核系统中,信号肽与SRP(信号识别颗粒)的结合亲和力直接决定了翻译后转运的比例,亲和力越高,成熟蛋白的累积产量越大。反之,若信号肽序列与宿主分泌系统不匹配,可能导致前体蛋白在胞质中积累并被蛋白酶降解,反而降低最终得率。因此,合理选择与宿主兼容、疏水性适中的信号肽,是优化蛋白表达量的首要策略。
Q2:信号肽能否增强蛋白的可溶性表达?
A:信号肽能显著增强蛋白的可溶性表达,但其效果依赖于目标蛋白特性和宿主系统。在细菌如大肠杆菌中,信号肽将蛋白导向氧化性的周质空间或直接分泌至培养基。周质空间内含有DsbA等二硫键异构酶,能帮助形成正确二硫键,这对于抗体片段、细胞因子等富含二硫键的蛋白尤为重要。相比胞质还原环境,信号肽介导的周质表达常使可溶性部分占比从不足10%提升至50%以上。
因此,实验前需要先通过软件(如SignalP、PrediSi)预测信号肽与宿主的适配性,并设置胞质表达对照,通过SDS-PAGE和Westernblot确认可溶性提升效果。
Q3:在纯化过程中,信号肽本身是否可以直接用作纯化标签?
A:信号肽本身很少直接用作纯化标签,因为其高度疏水的短肽序列缺乏特异性结合配体,无法像His标签或GST那样通过亲和层析捕获。但信号肽切割后留下的N端序列或工程化改造的信号肽可以间接辅助纯化。常见的实用策略有两种:
第一,将信号肽与常规亲和标签融合表达。例如,在信号肽与目标蛋白之间插入His标签或Strep标签。当信号肽引导分泌后,信号肽被信号肽酶切除,标签留在成熟蛋白N端,仍可用于亲和纯化。这种方式既利用了信号肽提升表达量和可溶性的优势,又保留了便捷纯化的能力。
第二,部分系统利用信号肽的“锚定”特性进行初始富集。例如在酵母中,使用α-因子信号肽融合目标蛋白后分泌至上清,上清液经超滤浓缩后可直接进入离子交换或疏水层析。信号肽的存在不会干扰纯化,因为它在分泌时已被移除。
总之,信号肽更适合作为“表达增强元件”而非直接纯化工具,与其结合成熟的亲和标签才能发挥最大效用。
Q4:原核与真核宿主中,不同信号肽对蛋白表达和纯化效果的影响有哪些差异?
A:原核与真核宿主的细胞结构、转运系统差异,导致信号肽的作用效果存在显著区别。在大肠杆菌等原核宿主中,常用PelB、OmpA等信号肽,主要引导蛋白转运至周质空间,操作简单、剪切效率高,可有效提升可溶性表达量,纯化时周质蛋白杂质少、提纯难度低,适合原核快速表达纯化体系。
在酵母、哺乳动物细胞等真核宿主中,常用SP、IgK等信号肽,可介导蛋白高效胞外分泌,大幅提升上清液中目标蛋白浓度。但真核信号肽序列更长、修饰更复杂,部分序列易出现剪切不完全的问题,会增加纯化后的蛋白均一性调控难度。整体而言,原核信号肽侧重提升纯化效率,真核信号肽侧重提升胞外表达量。
Q5:无信号肽胞内表达与信号肽介导的分泌表达,在蛋白纯化工艺上有哪些核心优劣区别?
A:无信号肽的胞内表达与信号肽介导的分泌表达,是重组蛋白制备的两种核心方式,在纯化工艺、产物质量、操作成本上差异显著。无信号肽表达的蛋白主要滞留于细胞质内,表达总量相对稳定,但胞内含有大量杂蛋白、核酸、内毒素与细胞碎片,纯化前期需经过破菌、除杂、脱毒等复杂工序,流程繁琐且产物损耗高。
信号肽介导的分泌表达可将目标蛋白富集于周质空间或胞外上清,杂质组分简单,无需复杂破菌步骤,能大幅简化纯化流程、降低工艺成本。同时该方式可减少蛋白胞内堆积引发的包涵体生成,提升可溶性蛋白占比。唯一劣势是部分信号肽分泌效率有限,极端条件下会轻微降低总表达量,需通过信号肽改造平衡产量与纯化难度。
- 联系我们
- 电话:400-621-6806
- 手机:13116038708(微信)
- 邮箱:info@kmdbioscience.com
- 地址:天津市武清开发区和畅路6号10号厂房108室
-

销售微信
-

官方公众号
-
 400-621-6806
400-621-6806 -
 3599163699
3599163699 -
 周一至周五 9:00-18:00
周一至周五 9:00-18:00 -
-
 0
0
-
