

Nature综述后的首次突破:David Baker团队实现从头设计蛋白质随机步行者!
- 2026-06-18
- 9
一.文献背景
2023年诺贝尔化学奖得主David Baker团队在Nature发表综述,系统阐述蛋白质从头设计技术从物理计算到AI生成的范式转变,及其在药物开发、纳米机器等领域的突破性进展。 该技术有望在5-10年内设计出攻克重大疾病和环保难题的蛋白质解决方案。1.Molecular walkers
文中提到的Molecular walkers,位于生物学,化学和物理学的迷人十字路口,能将能量转化为单向运动。天然分子马达,如动力蛋白,驱动蛋白和肌球蛋白,它们是生命活动所必需的,虽然已有详细研究,但我们对它们的理解仍然不完整。可编程和健壮的从头设计的蛋白质纳米机器人可以彻底改变从精密医学到材料的领域。尽管最近在从头蛋白质设计方面取得了显著的进展,但构建这样的功能系统仍然遥不可及。
2.研究现状
现有技术包括由天然DNA结合蛋白组装的蛋白质步行者,其可以在微流体和从头设计的转子蛋白的帮助下沿着DNA台阶进行移动。跟踪转子的运动被证明是具有挑战性的工作,设计一种随机蛋白质步行者,可以沿着很长的距离移动,这将标志着一个实质性的进步,加深我们对分子机器设计的理解,并为未来基于蛋白质的纳米器件奠定基础。
3.研究方向
分子机器具有巨大的潜力,静态单体和寡聚体蛋白质结构的设计已经取得了巨大的进展,但很少有动态蛋白质系统的设计。
基于以上现状,Baker团队2025年在预印本平台bioRxiv发表了题为"De-novo design of a random protein walker"的研究论文,首次实现了从头设计的蛋白质随机行走系统。这一突破标志着蛋白质设计从静态结构创造迈向动态功能构建的关键转折,为开发可编程的"蛋白质机器人"提供了全新的技术平台。

4.研究的优势
▶结合特异性:作者设计一个完全从头设计的蛋白质系统,由4到8英尺的步行者组成,这些步行者可以特异性地结合到具有立足点的设计蛋白质轨道上。
▶0消耗:这些步行者在不消耗燃料的前提下沿着轨道的扩散成为可能并表征其特征,从而探索多价性如何影响速度和持续合成能力。这个最小的系统提供了一个多功能的支架,外部能源可以在未来的设计中融入。
二.研究成果
1. 轨道的设计策略和结构特征
作者推断至少需要三个组件来构建随机步行者:长蛋白质纤维作为轨道,对称蛋白质支架作为步行者,以及行为良好的可逆异二聚体作为脚和立足点,形成关键界面,使步行者能够沿着轨道前进。
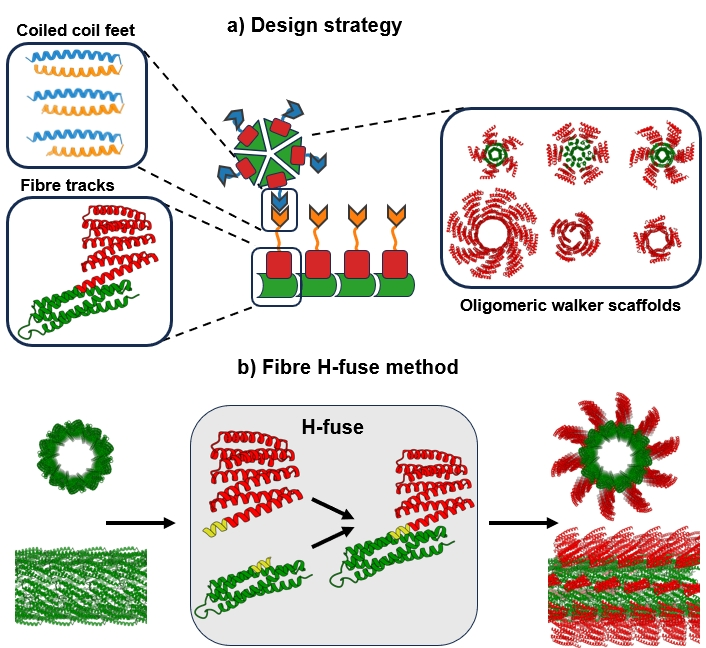
图1 随机步行者系统的基本部件
由球状单体自组装而成的蛋白质纤维已经被成功地设计出来,但是它们太短了,无法用全内反射荧光(TIRF)显微镜(单分子追踪实验的关键技术)清楚地分辨出纤维。为了解决这个限制,作者的目标是设计具有更大直径的纤维,以增加它们的刚性,进一步提高纤维长度和结构稳定性。
轨道的核心就是通过对接蛋白质支架和重新设计接口来设计的,作者开发了Fibre H-fuse,这是一种将任意蛋白质刚性连接到纤维组装体上的新方法。其中两种蛋白质-HA 4和HA 13-都以可溶性形式表达并形成纤维,这一点得到了共聚焦显微镜的证实,通过分辨率为3.3 nm的cryoEM进一步表征HA 13的结构,该结构揭示了具有10 nm内径和连接的DHR之间的4.7 nm纵向间距的纤维。
除了便于电子显微镜检查外,DHR的融合还提供了与纤维界面的额外间隔,减少了在为步行者添加立足点时扰乱纤维组装的机会。
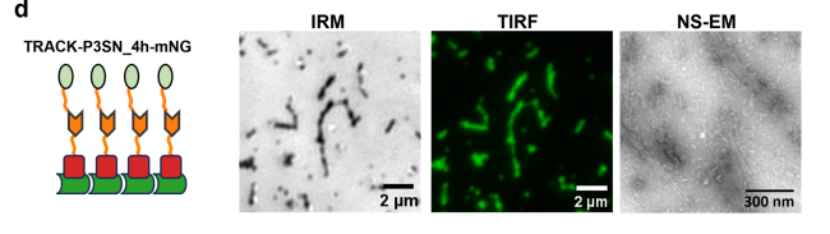
图2-1 轨道的结构特征
2. 异二聚体足/立足点的选择
为了完成跟踪,需要能够结合和解结合的异二聚体原体作为纤维上的立足点。用较短的异二聚体四螺旋束mALb 811进行的初步测试表明,立足点的存在会干扰纤维组装。
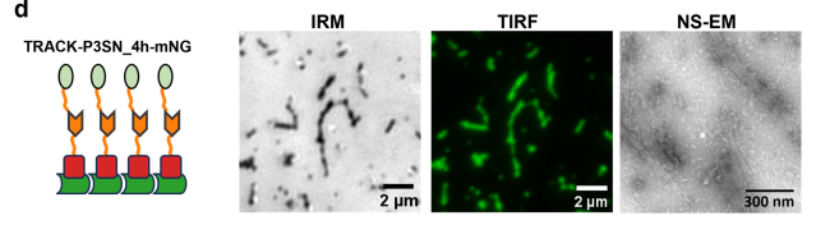
图2-2 轨道的结构特征
3. 步行者设计
作者通过柔性六残基接头将P4 SN卷曲螺旋足连接到支架上; P4 SN特异性结合到纤维上展示的互补P3 SN卷曲螺旋足。
所有六个步行者构建体在大肠杆菌中成功表达并从可溶性级分中纯化。使用两种尺寸排阻法验证了寡聚体组装状态,分别是在微摩尔和纳摩尔浓度下进行的色谱-多角度光散射(SEC-MALS)和质量光度法(MP),这两种技术都证实了所设计的组装体的预期分子量和低聚状态。

图3 设计的步行者寡聚体的生物物理表征和结构验证
4. 轨道上步行者的单分子跟踪
为了研究步行者沿着轨道的动力学,我们使用TIRF显微镜和荧光显微镜,使用TIRF在绿色通道中可视化跟踪,实现无标签成像。用ATTO 643荧光标记步行者,并通过TIRF以4 Hz的帧速率在远红通道中成像。通过使用Picasso 17拟合荧光强度分布的中心来定位步行者位置。从步行者轨迹中,提取了步长的分布,并将其拟合为二维高斯函数。
测量了具有不同数量的脚沿着其余足迹的步行者的扩散,出现了两个主要趋势:与具有较低结合亲和力(P3SN_3.5 h)的那些相比,步行者在具有较高结合亲和力足点(P3SN_4 h)的轨道上移动得更快,并且扩散速率随着步行者效价而增加。
为了评估附着在轨道上的mNeonGreen是否会影响步行者的移动性,测量了WALKER-C4和WALKER-C8在缺乏荧光蛋白的轨道上的扩散。在有和没有mNeonGreen的轨道上的扩散速率相当,表明柔性拴系融合体的存在不会对步行者一旦绑定到轨道上的移动产生可测量的影响。
另外还评估了温度、粘度和盐浓度对WALKER-C8在缺乏mNeonGreen的轨道上移动的影响。(25°C时为1.695cP)不会使步行者减速,在50%蔗糖时,其在两条轨道上的扩散速率下降约2倍在500 mM时,扩散速率也下降了约1.5倍,这很可能是由于足和立足点之间的亲和力降低。相反,当温度从25 ℃升高到45 ℃时,它们的扩散速率增加约1.5倍。
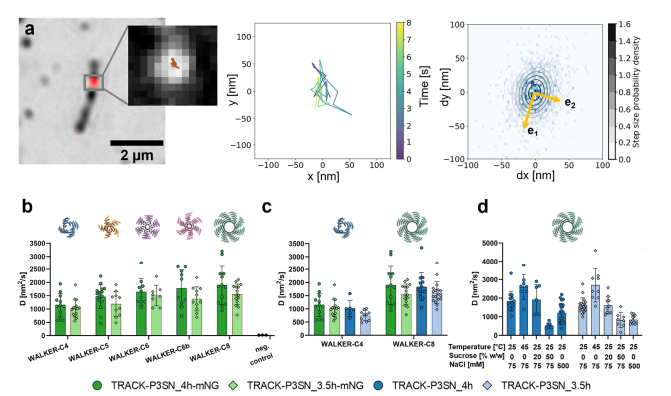
图4 工程随机蛋白质步行者的单分子追踪和扩散分析
有关蛋白质重组设计的研究课题热度不断,卡梅德生物可以提供多种蛋白表达,重组蛋白设计,重组蛋白制备及蛋白纯化等定制化服务服务,满足客户在不同实验阶段的需求。
[1] https://doi.org/10.1101/2025.09.29.677966
常见问题汇总
Q1:设计阶段的那些问题会导致蛋白表达失败?
A1:
密码子偏好性不匹配:如果外源基因含有宿主罕见的密码子,会严重拖慢翻译速度。此时可对基因序列进行密码子优化,或使用 Rosetta 等补充稀有 tRNA 的专用菌株。
信号肽问题:将真核蛋白的全长序列(含信号肽)直接用于原核表达,会导致错误折叠。克隆前应切除信号肽。
标签选择不当:N 端标签可能因信号肽而被切除。对于易降解的蛋白,可将融合标签放在 N 端;若为分泌蛋白,则推荐 C 端标签。
Q2:表达阶段的那些问题会导致蛋白表达失败?
A2:
表达量低:这是最常见的困境。除了对基因和表达系统进行优化,还应调整诱导条件(时机、温度、诱导剂浓度),并尝试不同浓度的裂解液。
形成包涵体:蛋白过量或快速表达,来不及正确折叠,就会聚集成无活性的“包涵体”。可通过低温慢诱导(如25℃)、使用弱启动子或在培养基中加入山梨醇等渗透调节剂来改善。对于原核宿主,也可更换为 Origami 等促进二硫键形成的菌株。
蛋白降解:蛋白酶是细胞内的“清道夫”,会攻击外源蛋白。核心解决思路是“减少暴露”和“抑制活性”。比如缩短诱导后的培养时间、在裂解缓冲液中加入蛋白酶抑制剂、更换缺乏蛋白酶基因(如 lon, ompT)的宿主菌株。
翻译后修饰不足:蛋白功能的实现,常依赖糖基化、磷酸化等“后期修饰”。如果“不足”,很可能是宿主能力不足。对此,可以从简单的大肠杆菌系统,升级到酵母、昆虫甚至哺乳动物细胞系统来解决问题。
Q3:重组蛋白制备流程?
A3:基因克隆与载体构建:将目的基因插入表达载体,配备启动子、标签(如 His 标签)等元件。
宿主转化与表达筛选:将载体导入宿主细胞,通过小规模培养筛选阳性克隆。
扩大培养与诱导表达:筛选成功后,放大培养并优化诱导时间、温度、诱导剂浓度等参数。
细胞裂解与样品澄清:收集细胞,用超声或高压匀浆等方式裂解,离心后获得含目标蛋白的上清液。
蛋白纯化:利用亲和层析、离子交换层析等多步纯化策略,获得高纯度蛋白。
Q4:表达系统的选择原则及其优势?
A4:
表达系统 | 优点 | 缺点 | 最佳适用场景 |
大肠杆菌 (E. coli) | |||
酵母 | 兼具原核的操作性和真核的折叠修饰能力,可分泌表达 | 存在过度糖基化问题,糖型与人不同 | |
昆虫细胞 | |||
哺乳动物细胞 | 药物研发、抗体生产等需完全人源化蛋白的应用,被视为“金标准” |
Q5:蛋白纯化后续可能会出现的问题?
A5:
纯化后蛋白纯度低:常因His标签未有效暴露或洗涤不充分。可通过在缓冲液中添加低浓度咪唑、甘油或改变pH来改善。
蛋白溶解性差:原因包括溶液pH接近其等电点导致沉淀,或浓度过高发生聚集。应优化缓冲液配方(pH、离子强度),降低蛋白浓度,并在低浓度工作液中加入BSA等载体蛋白提高稳定性。
蛋白活性低或无活性:常因复性失败或储存不当。需优化包涵体复性条件,或立即分装冻存以避免反复冻融。
- 联系我们
- 电话:400-621-6806
- 手机:13116038708(微信)
- 邮箱:info@kmdbioscience.com
- 地址:天津市武清区国际企业社区A5号楼
-

销售微信
-

官方公众号
-
 400-621-6806
400-621-6806 -
 3599163699
3599163699 -
 周一至周五 9:00-18:00
周一至周五 9:00-18:00 -
-
 0
0
-
