

合作成果|激活还是抑制?揭开微生物防御素对新冠的“双面人生”
- 2026-05-22
- 97
| 期刊 | 《Fontiersin Microbiology》 |
| 题目 | 《Enhancement of SARS-CoV-2 receptor-binding domain activity by two microbial defensins》 |
| 影响因子 | 6.6(2023年) |
| 单位 | 中国科学院动物研究所、农业虫害鼠害综合治理研究国家重点实验室 |
| 引用卡梅德(KMD Bioscience)产品/服务 | 该研究中的α变异株RBD(产品编号:COV390)、β变异株RBD(产品编号:COV380)、γ变异株RBD(产品编号:COV381)、ζ变异株RBD(产品编号:COV391)和hACE2蛋白(产品编号:KMPH1835)由天津卡梅德生物(KMD Bioscience)提供,主要用于表面等离子共振SPR分析,以精确测量两种防御素肽(AMSIN和emorisin)与不同SARS-CoV-2变异株RBD的结合亲和力,为病毒相关研究提供基础与保障。 |

防御素是微生物先天免疫的重要效应分子,也是探索病毒蛋白调控机制的关键研究靶点,SARS-CoV-2通过刺突蛋白的受体结合域(RBD)与人体ACE2结合实现细胞入侵,针对RBD的结合分子研究对解析病毒侵染机制、开发相关分子工具具有重要理论与应用价值。
2023年6月,中国科学院动物研究所朱顺义带领的研究团队首次发现可与SARS-CoV-2受体结合域(RBD)结合的AMSIN和emorisin两种微生物防御素。该研究成果拓宽新冠病毒RBD变构调控的分子机制研究,为新冠病毒诊断试剂研发开辟全新思路与方向。
一、研究概述
SARS-CoV-2病毒通过刺突蛋白的受体结合域与人体细胞表面的血管紧张素转换酶2结合完成细胞侵染,研发靶向该受体结合域的结合分子对病毒相关研究和抗病毒药物开发具备重要价值。不同于以往发现的抑制性结合分子,分离自放线菌的AMSIN与源自真菌Emergomycesorientalis的emorisin能以7.6–1450nM的中高亲和力结合野生型及α、β、γ、δ、ζ等多种变异株的受体结合域,并通过正变构效应提升受体结合域与血管紧张素转换酶2的结合活性。
本研究整合结构学生物化学与计算生物学技术方法,阐明这一独特功能的分子机制:两种防御素均拥有保守的二元结构基序,依靠阳离子-π相互作用启动受体结合域的变构信号传递,最终实现对血管紧张素转换酶2结合位点构象的精准调控。此外,固定于CM5传感器芯片的AMSIN具备优异稳定性,在人血清环境中依旧可与受体结合域高效结合,显现出作为新冠病毒抗原诊断工具的应用前景。这项研究拓宽对蛋白质变构调控的认知边界,也为新冠病毒相关的基础研究与应用开发打造全新的分子工具。
二、Emorisin的鉴定与天然结构特征验证
通过序列比对、结构建模及理化验证,明确了emorisin与AMSIN的序列特征、保守功能基序及天然构象,结果表明二者虽序列相似度仅40%,但均含由阳离子残基与芳香族残基构成的保守二元结构基序,且化学合成的emorisin可成功实现氧化复性,形成纯度高、分子量匹配的天然CSαβ型防御素构象,为后续探究其与SARS-CoV-2RBD的相互作用奠定了结构基础。
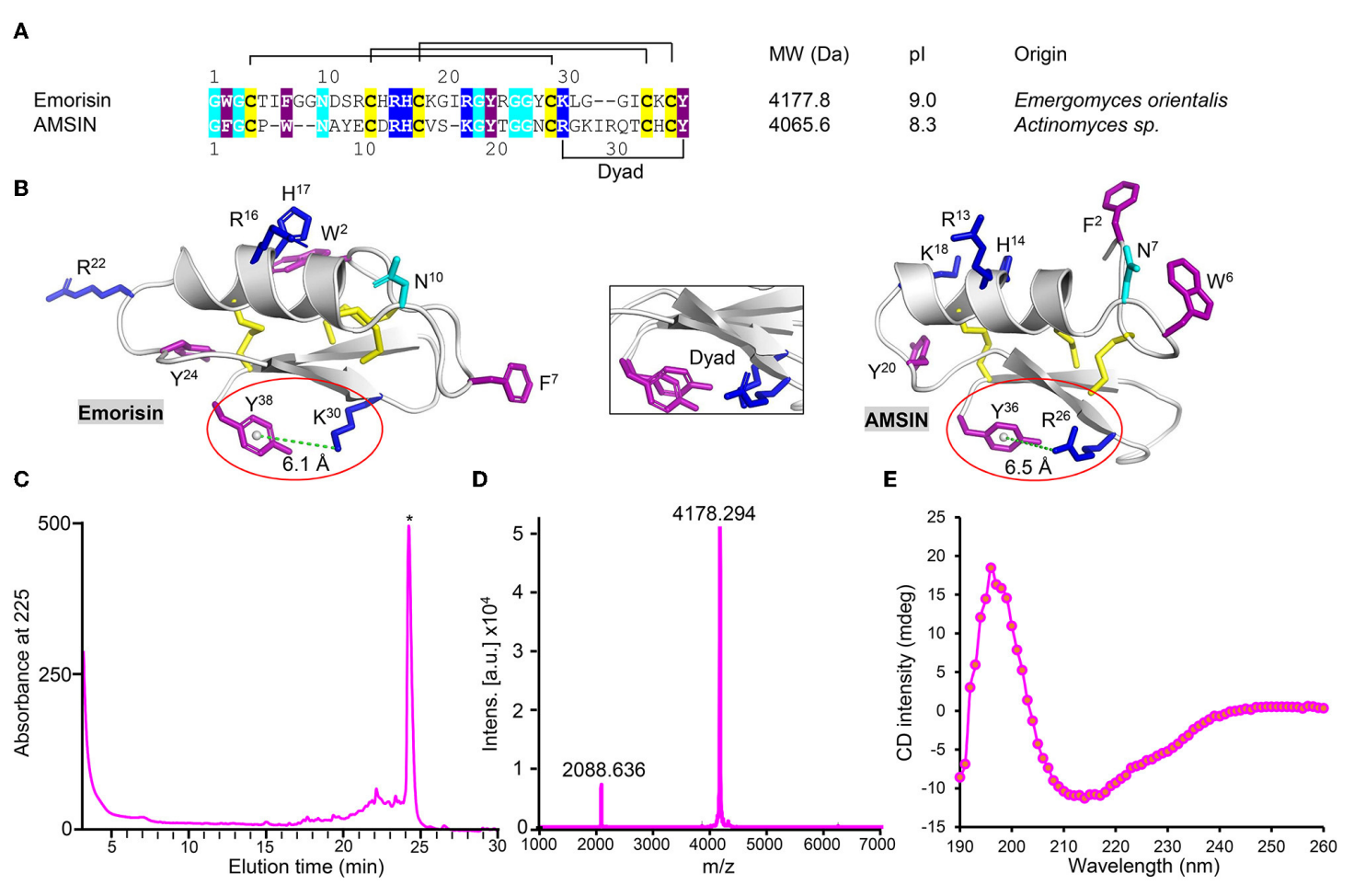
图1 emorisin鉴定与结构特征
三、AMSIN与Emorisin对不同RBD的结合动力学特征
SPR实验结果表明,AMSIN可与野生型及α、β、γ、δ、ζ等多种变异株的RBD及Omicron变异株的oSpike蛋白发生特异性结合,结合具备明显的浓度依赖性且解离过程稳定,对αRBD和oSpike蛋白展现出高亲和力,对不同RBD均有结合能力且结合过程快速稳定,体现出广泛的RBD识别特性。
同时,emorisin与RBD的结合曲线呈现出和AMSIN相似的特征,且对WTRBD、βRBD和δRBD的结合亲和力优于AMSIN,不过emorisin仅能与特定类型的RBD发生结合,对α、γ、ζRBD未检测到结合信号,说明其对RBD的识别具有选择性,结合位点与AMSIN存在差异。
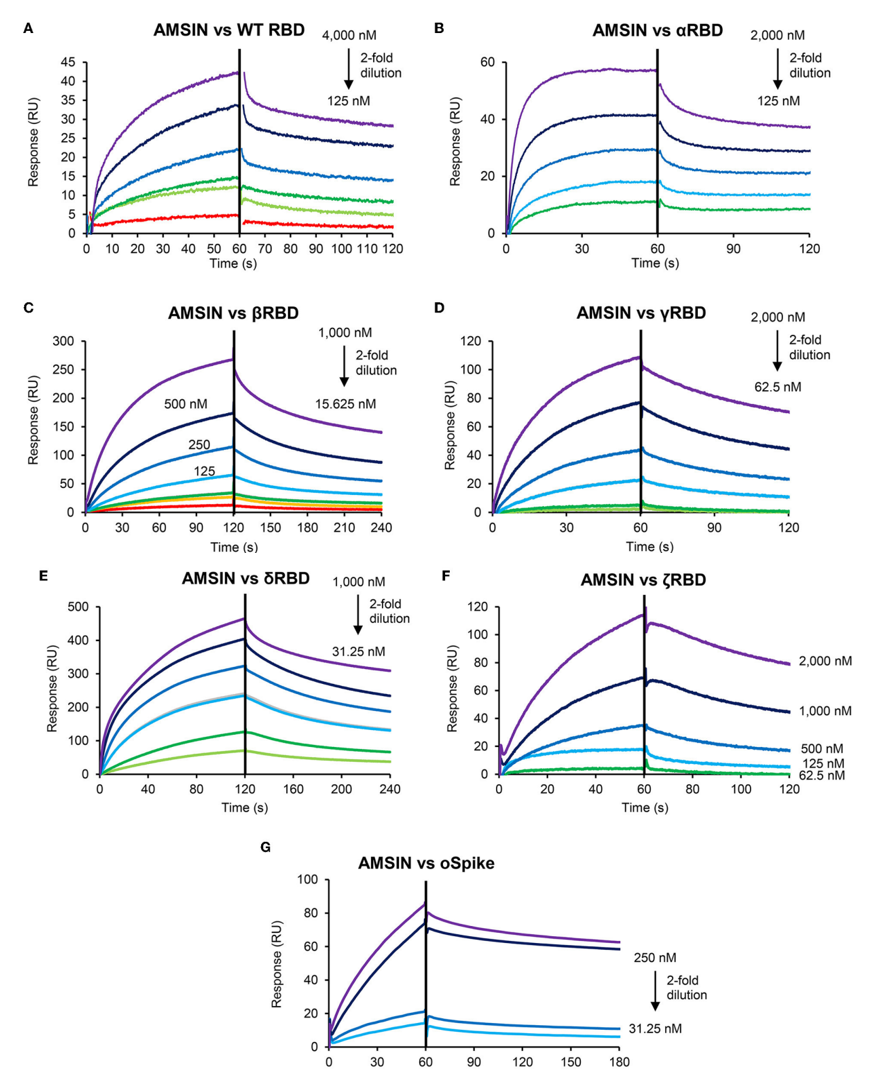
图2 AMSIN与各RBD变异株的SPR传感图
图3 emorisin与RBD的SPR传感图
四、防御素增强RBD与ACE2结合的正变构效应
研究结果表明,AMSIN与emorisin均能以浓度依赖性方式显著增强WTRBD与ACE2的结合活性,其中AMSIN的激活效应经SPR竞争实验和ELISA实验双重验证,emorisin的激活效率随浓度提升而增强且存在浓度阈值,二者均通过非竞争结合的方式发挥正变构调控作用。
野生型RBD内存在与ACE2结合位点空间相连的变构相关残基簇,该残基簇是变构信号传递的核心通路,在各变异株中保守性极高,其中AMSIN以自身Arg-26与RBD的Phe-342构建阳离子-π相互作用。该相互作用作为诱导RBD发生变构的核心信号位点,可借助保守残基簇将信号传导至ACE2结合位点,促使该区域构象改变,最终提升RBD与ACE2的结合活性。
图4 不同实验证实AMSIN具有激活作用
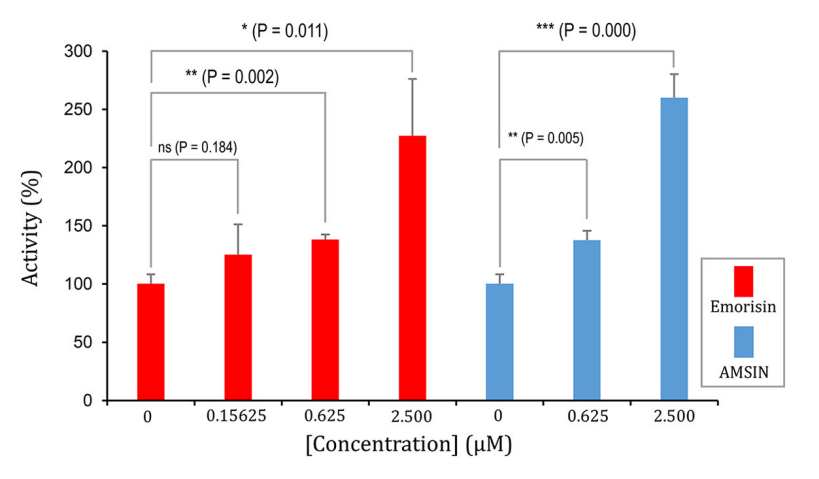
图5 ELISA实验验证emorisin具有激活效应
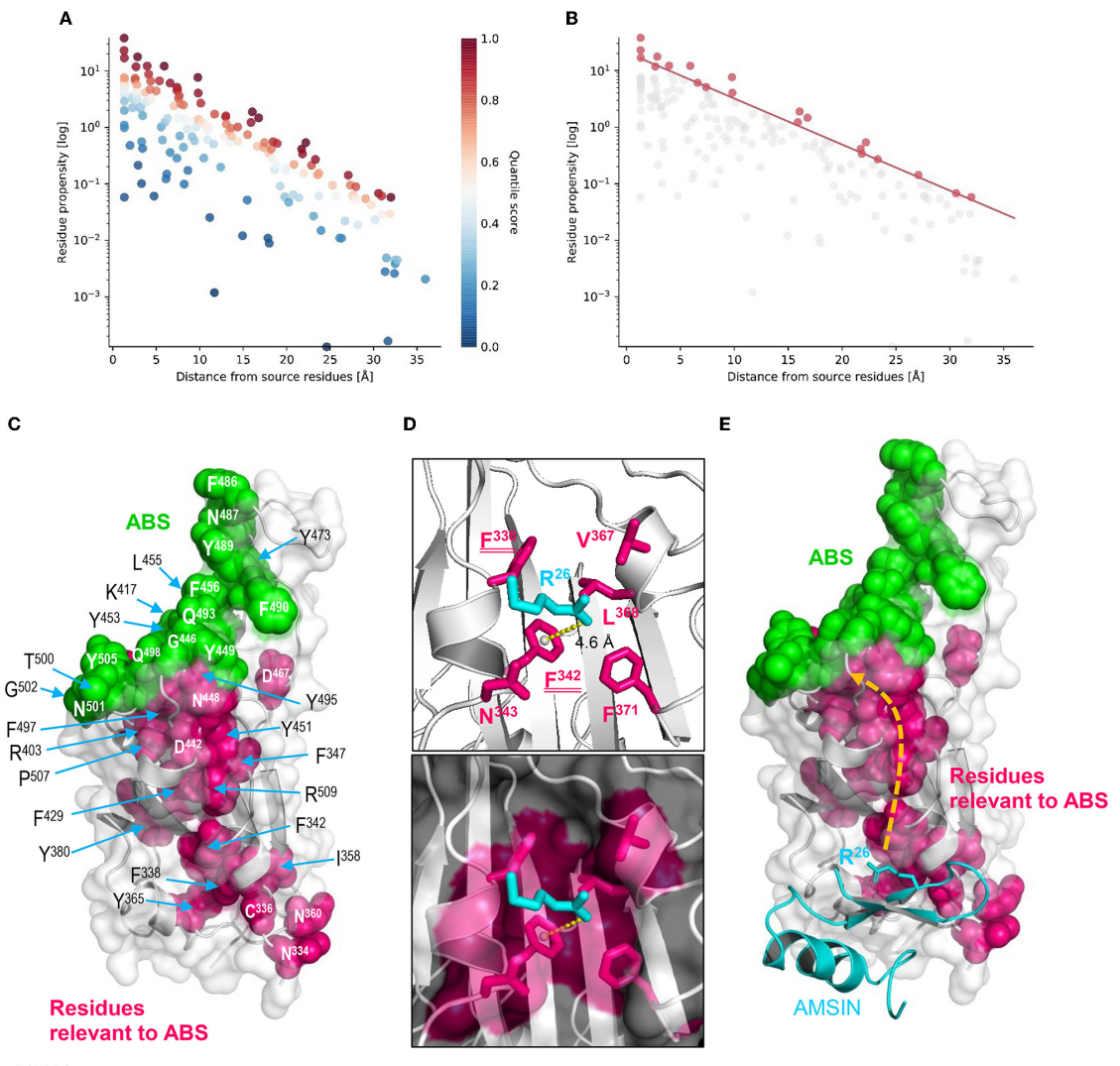
图6 RBD中由AMSIN诱发的配体调节作用的结构基础
五、AMSIN用于新冠病毒诊断的潜力验证
选取δRBD作为靶标,在1%人血清环境中检测AMSIN与δRBD的结合作用,SPR结果显示人血清的存在未干扰二者的结合过程。SDS-PAGE电泳证实,固定于CM5芯片的AMSIN拥有优异的抗血清干扰特性,可将其开发为新冠病毒抗原检测试剂,进一步印证AMSIN在新冠病毒诊断领域的潜在应用前景。
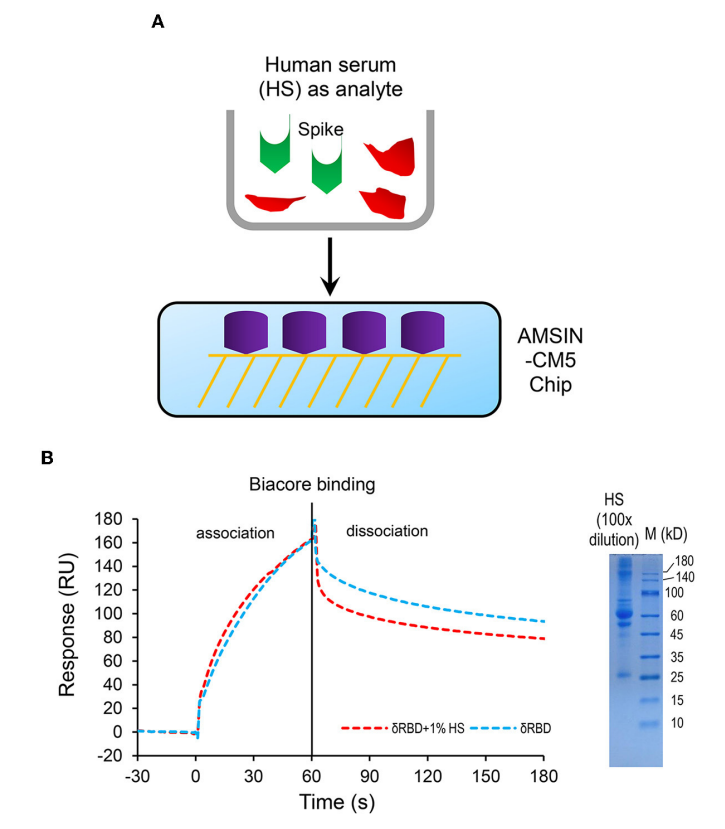
图7 人血清不会干扰δRBD与共价结合在CM5上的AMSIN之间的结合
本研究依托系统的结构生物学与生物化学解析手段,首次筛选得到具备正变构激活作用的SARS-CoV-2RBD结合分子,颠覆此前防御素类分子以抑制性作用为主的固有认知。其中两种防御素的特有作用方式为探究RBD变构调控机制提供全新的分子探针,二者兼具的高亲和力、广谱结合性与血清稳定性等特性为新冠病毒相关基础研究和应用开发开拓全新路径。
该研究中的α变异株RBD(产品编号:COV390)、β变异株RBD(产品编号:COV380)、γ变异株RBD(产品编号:COV381)、ζ变异株RBD(产品编号:COV391)和hACE2(产品编号:KMPH1835)购自KMDBioscience(中国天津),主要用于表面等离子共振SPR分析,以精确测量两种防御素肽(AMSIN和emorisin)与不同SARS-CoV-2变异株RBD的结合亲和力,为病毒相关研究提供基础与保障。
卡梅德生物(KMD Bioscience)依托成熟的噬菌体展示平台,可构建高达10^13/mL容量的随机肽库及环肽库,同时提供多类型预制肽库及固相、液相、细胞、磁珠等多种筛选方法。经蛋白互作技术(BLI、SPR)分析,筛选出的靶向肽段亲和力可达nM级,实现从高通量筛选到精准表征的一体化解决方案,助力您的科研成果加速转化。
[1]GaoB,ZhuS.EnhancementofSARS-CoV-2receptor-bindingdomainactivitybytwomicrobialdefensins.FrontMicrobiol.2023;14:1195156.
Q1:微生物防御素对新冠病毒的“激活”与“抑制”双重作用具体表现是什么?
A:微生物防御素对新冠病毒的双重作用,核心源于其与病毒蛋白、宿主细胞的不同互作模式。
抑制作用主要体现在阻断病毒入侵与复制:部分防御素(如人β-防御素4、肠道防御素5)可特异性结合新冠病毒刺突蛋白(S蛋白)的受体结合域(RBD),竞争性抑制RBD与宿主ACE2受体的结合,同时破坏病毒包膜完整性,阻止病毒内化进入宿主细胞,且能抑制病毒基因组复制与装配,降低病毒载量。
激活作用则表现为异常调控宿主免疫反应:部分微生物防御素可结合RBD的非ACE2结合位点,通过别构效应增强RBD与ACE2的亲和力,促进病毒入侵,或过度激活 Toll 样受体(TLRs)通路,引发细胞因子风暴,加重肺部炎症损伤,不利于病情恢复。这种双面性与防御素的种类、浓度、作用靶点及宿主免疫状态密切相关,也是当前科研探索的核心重点,而蛋白互作分析技术是解析该机制的关键手段。
Q2:如何通过多肽筛选技术挖掘具有抗新冠活性的微生物防御素?
A:多肽筛选技术是高效挖掘抗新冠微生物防御素的核心路径,需结合多平台技术实现精准筛选。
首先,基于微生物基因组数据库,通过生物信息学分析预测潜在防御素序列,利用固相合成技术构建高纯度防御素多肽文库,可引入非天然氨基酸优化序列,提升文库质量与多样性。
其次,采用高通量筛选策略,以新冠病毒S蛋白、ACE2受体为靶点,通过细胞水平筛选,初步筛选出能抑制病毒复制的候选多肽,同时借助噬菌体展示技术富集高亲和力结合肽,排除假阳性结果。
最后,通过二次筛选验证候选防御素的活性,结合蛋白互作分析(SPR、BLI)测定其与靶标蛋白的结合亲和力,筛选出纳摩尔级高活性防御素,同时排除具有激活作用的序列。
Q3:微生物防御素发挥抗新冠作用的分子机制主要有哪些??
A:微生物防御素发挥抗新冠作用的分子机制主要分为直接抗病毒机制与间接免疫调节机制,二者协同发挥作用。
直接抗病毒机制核心是靶向病毒结构蛋白:一是通过阳离子特性结合新冠病毒包膜,破坏包膜的电化学梯度,导致病毒颗粒裂解,阻止病毒感染宿主细胞;二是特异性结合S蛋白RBD域,阻断RBD与ACE2受体的相互作用,抑制病毒吸附与内化,部分防御素(如HD5)可通过“掩盖”ACE2受体,进一步阻断病毒入侵路径。
间接免疫调节机制则是通过调控宿主先天免疫反应,适度激活免疫细胞(如巨噬细胞、中性粒细胞),促进抗病毒细胞因子分泌,增强机体清除病毒的能力,同时抑制促炎因子过度释放,避免炎症损伤。此外,部分防御素可通过抑制病毒基因组复制、干扰病毒装配释放,进一步发挥抗病毒作用。这些机制的解析,需依托多肽合成、蛋白互作分析(SPR/BLI)等技术,精准验证关键作用位点与互作模式。
Q4:防御素多肽序列极短且结构复杂,化学合成时需要注意什么?
A:防御素的特征是富含半胱氨酸,能够形成复杂的分子内二硫键,这对维持其抗病毒活性至关重要,但也给化学合成带来了挑战。目前,最常用的是Fmoc固相合成法。在合成防御素时,需要注意以下几点:
(1)序列长度与修饰:防御素通常在30-50个氨基酸左右,合成难度适中,但若需要引入特定的荧光标记或生物素标记以进行互作分析,需要在合成设计阶段就规划好修饰位点和策略。
(2)二硫键的正确配对:采用“选择性半胱氨酸保护”策略,即使用不同的保护基团保护不同位置的半胱氨酸,在肽链合成完毕后,分步脱保护并氧化,从而精准控制二硫键的连接方式。
(3)纯化与质控:需要使用高效液相色谱法将目标肽段与合成杂质、错配异构体分离开,并通过质谱法精确验证分子量和二硫键连接方式的正确性,确保用于后续实验的防御素多肽具有正确的结构和活性。
Q5:除了直接结合病毒蛋白,防御素还有哪些抗病毒机制可以研究?
A:防御素的抗病毒作用机制是多样化的,除了直接与新冠病毒刺突蛋白结合外,还存在多种间接或辅助性的“赋能”机制值得深入探索。
(1)免疫调节作用:防御素作为先天性免疫的效应分子,可以招募和激活其他免疫细胞(如T细胞、树突状细胞),调节炎症因子的分泌,从而帮助宿主建立更强大的抗病毒免疫环境。
(2)破坏病毒包膜:防御素的阳离子特性使其能够与某些病毒的脂质包膜发生静电相互作用,插入并破坏包膜的完整性,导致病毒颗粒失活。对于有包膜的新冠病毒,这同样可能是一种抑制机制。
(3)病毒感染:防御素可能影响主要组织相容性复合体分子对病毒肽段的提呈,从而改变宿主免疫系统识别和清除感染细胞的能力。这些研究方向的深入,将更全面地揭示防御素在机体抗病毒防御中的“赋能”角色。
- 联系我们
- 电话:400-621-6806
- 手机:13116038708(微信)
- 邮箱:info@kmdbioscience.com
- 地址:天津市武清区国际企业社区A5号楼
-

销售微信
-

官方公众号
-
 400-621-6806
400-621-6806 -
 3599163699
3599163699 -
 周一至周五 9:00-18:00
周一至周五 9:00-18:00 -
-
 0
0
-
