

疫苗效果又双叒打折?可能是E484位点这个“免疫原性引擎”没调对
- 2026-06-05
- 38
| 期刊: | 《Viruses》 |
| 文章标题: | 《Enhancing the Immunogenicity of RBD Protein Variantsthrough Amino Acid E484 Mutation in SARS-CoV-2》 |
| 影响因子: | 4.8 |
| 客户单位: | 南方医科大学、广东省热带病研究重点实验室、南方医科大学南方医院 |
| 引用卡梅德产品/技术服务: | 该研究中使用的重组人源抗SARS-CoV-2 S1 RBD单克隆抗体购自卡梅德生物(KMD Bioscience,中国天津),提供包含产品编号MAV101、MAV102、MAV103在内的三款不同类型核心试剂,主要用于Western blot抗原性鉴定与ELISA抗体效价分析,精准验证RBD突变体的体外抗原活性与体内免疫原性,为新冠病毒RBD功能研究及疫苗抗原筛选提供关键实验支撑。 |
新型冠状病毒SARS-CoV-2持续变异导致刺突蛋白受体结合域(RBD)高频突变,不仅增强病毒传播与免疫逃逸能力,也降低现有疫苗保护效果与抗体检测适用性。RBD是介导病毒结合宿主ACE2受体入侵的核心区域,更是重组蛋白疫苗、新冠抗体检测试剂盒开发的关键抗原靶点,筛选高免疫原性、广谱识别的RBD突变体,对打造适配变异株的抗疫工具具有核心价值。

2022年9月,南方医科大学研究团队在《Viruses》发表研究成果,通过大肠杆菌表达系统制备多种SARS-CoV-2 RBD突变体,证实E484位点氨基酸突变可显著增强RBD蛋白免疫原性,为广谱新冠疫苗设计与多变异株适配的抗体检测试剂开发提供关键候选靶点与技术支撑。
一、研究概述
SARS-CoV-2依靠S蛋白RBD与宿主细胞ACE2受体结合完成侵染,RBD上K417、L452、E484、N501为高频突变位点,其中E484位点存在E484K、E484Q、E484A等多种突变,广泛分布于Beta、Gamma、Kappa、Omicron等主流变异株,直接影响病毒侵染能力与免疫逃逸特性。
本研究以野生型RBD为模板,构建9种RBD突变体(6种单点突变、1种双点突变、2种三点突变),采用大肠杆菌原核表达系统制备蛋白,经包涵体变性复性、亲和纯化获得活性蛋白;通过Western blot验证体外抗原性,以小鼠免疫实验系统评价体内免疫原性,最终筛选出E484K、E484Q等高免疫原性突变体,明确E484位点对RBD功能与免疫原性的关键调控作用,同时验证大肠杆菌表达RBD可作为疫苗研发与抗体检测工具开发的高效载体。
二、RBD突变体的原核表达与纯化制备
本研究通过反向PCR技术构建了9种靶标突变体,包括6种单点突变体、1种双点突变体和2种三点突变体。经原核表达与蛋白纯化获得分子量约30kDa的活性重组RBD蛋白,采用亲和层析结合超滤去除内毒素后所有突变体内毒素含量均低于2.0EU/mL,各项指标符合动物实验与后续应用要求。
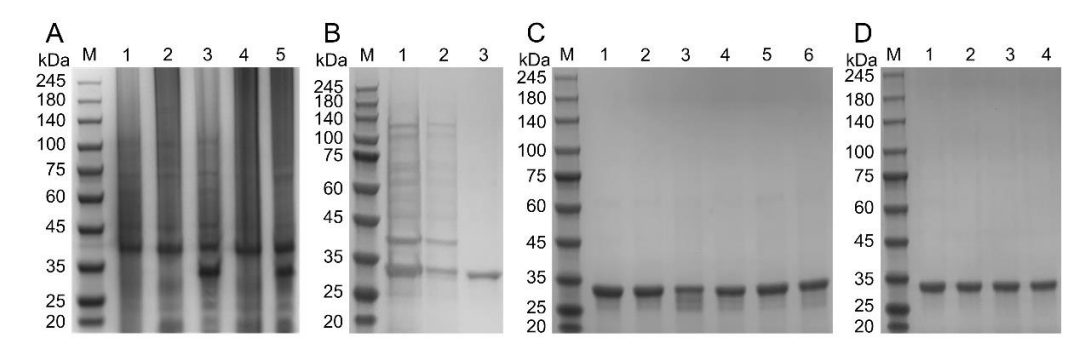
图1 RBD蛋白突变体的表达与纯化
三、RBD突变体具备强体外抗原性
采用Western blot免疫印迹检测纯化野生型与各突变型RBD蛋白的生物活性,其中野生型及9种突变体蛋白均在30kDa位置出现清晰的特异性条带,证实全部RBD突变体均具有良好的体外抗原性且抗原活性稳定可靠,可用于小鼠体内免疫原性评价等后续关键实验。
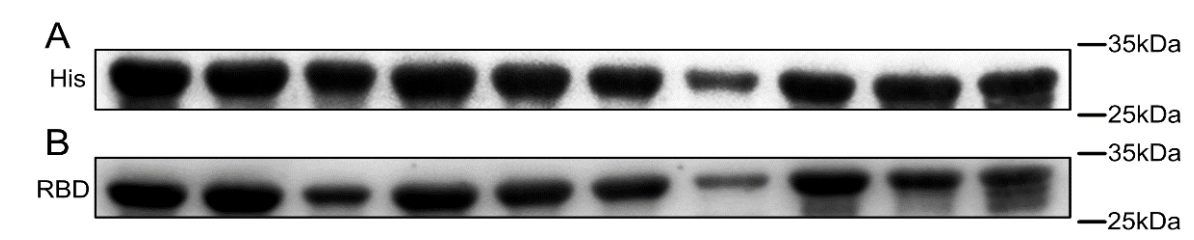
图2 RBD蛋白突变体的蛋白质印迹分析
四、E484位点突变是提升RBD免疫原性的核心关键
利用RBD蛋白免疫BALB/c小鼠,ELISA检测结果显示,免疫前所有小鼠血清均未检测到抗RBD抗体,免疫后第13天野生型与全部突变体均成功诱导产生特异性RBD抗体,第25天抗体水平大幅升高,不同突变体诱导的抗体效价呈现显著差异。其中E484K、E484Q、K417T‑E484K‑N501Y、K417N‑E484K‑N501Y诱导的抗体滴度分别为野生型RBD的4.7倍、4.8倍、6.8倍、5.0倍,免疫原性显著优于野生型;K417N、L452R、L452R‑E484Q组滴度约为野生型的2倍;K417T、N501Y单点突变与野生型无显著差异,该结果直接证明E484位点突变是提升RBD免疫原性的核心关键。
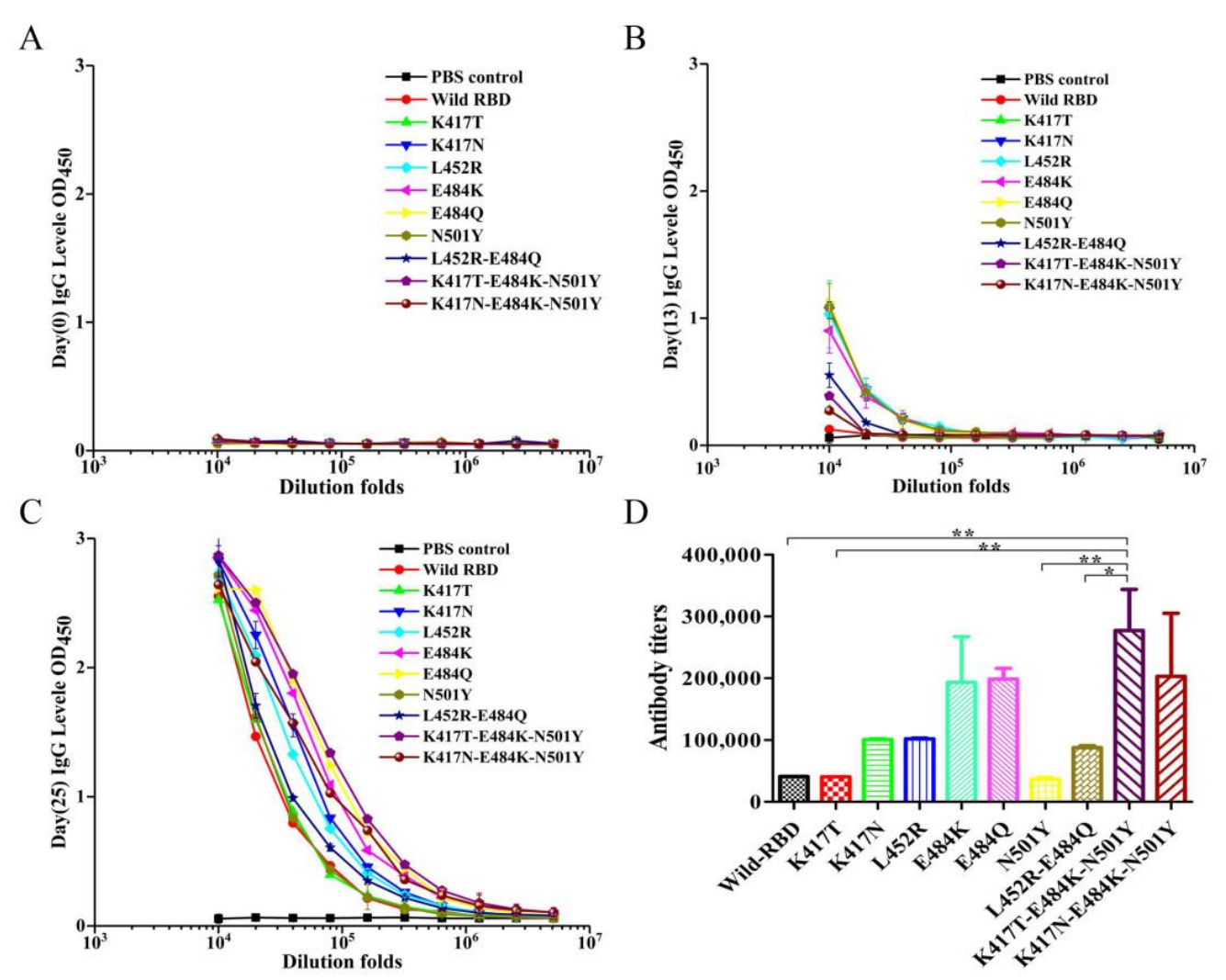
图3 通过酶联免疫吸附测定法检测小鼠体内针对新冠病毒 SARS-CoV-2 RBD 的抗体水平
五、E484位点对RBD广谱抗原性的关键调控作用
本研究分别以E484K、E484Q、K417T‑E484K‑N501Y、K417N‑E484K‑N501Y作为包被抗原,检测免疫小鼠血清的交叉结合效价。实验结果显示,E484K抗原可高效识别所有免疫组血清抗体,与野生型及多种突变体诱导的抗体结合能力优异,通用性突出;E484Q抗原诱导的抗体对广谱RBD突变体均呈现高结合效价,交叉识别能力极强;而两种三点突变体抗原仅对E484K、E484Q免疫组的抗体识别效果较好,对其他组别无显著优势,不适宜作为广谱检测抗原。该研究同时证实,RBD与ACE2的结合亲和力并不直接决定其免疫原性,这一重要发现为新冠疫苗的抗原设计与优化提供了全新理论依据。
图4 全面分析RBD变异蛋白的抗原性
本研究成功构建并验证9种新型冠状病毒SARS-CoV-2 RBD突变体的体外抗原性和体内免疫效果,并证实E484位点氨基酸改变可显著提升RBD蛋白免疫原性。研究不仅完善了新冠病毒RBD突变体功能调控机制的相关认识,而且为广谱重组蛋白疫苗开发和多变异株抗体检测试剂研制提供了关键抗原靶点实验数据,对提升病毒变异应对能力和健全新冠防控检测体系具有重要理论价值与应用前景。
该研究中使用的重组人源抗SARS-CoV-2 S1 RBD单克隆抗体购自卡梅德生物(KMD Bioscience,中国天津),提供包含产品编号MAV101、MAV102、MAV103在内的三款不同类型核心试剂,主要用于Western blot抗原性鉴定与ELISA抗体效价分析,精准验证RBD突变体的体外抗原活性与体内免疫原性,为新冠病毒RBD功能研究及疫苗抗原筛选提供关键实验支撑。
卡梅德生物依托成熟的抗体研发技术平台,构建覆盖杂交瘤、噬菌体展示、SingleB细胞筛选的多物种抗体发现平台,可开发鼠源、兔源、人源及羊驼VHH纳米抗体等多类型抗体,提供从单克隆/多克隆抗体制备到抗体人源化、亲和力成熟、双特异性抗体构建的一体化解决方案。同时,整合BLI、SPR亲和力分析与流式细胞术、ELISA特异性验证技术,筛选抗体亲和力可达pM级,为科研成果产业化提供坚实技术支撑。
[1] Zhang Z, Wan X, Li X, Cai S, Wan C. Enhancing the Immunogenicity of RBD Protein Variants through Amino Acid E484 Mutation in SARS-CoV-2. Viruses. 2022;14(9):2020.
常见问题汇总
Q1:为什么SARS-CoV-2的RBD区域是疫苗和检测试剂开发的关键靶点?
A:SARS-CoV-2的刺突蛋白(S蛋白)受体结合域(RBD)是病毒识别并结合宿主细胞ACE2受体的核心功能区。RBD通过构象变化与ACE2高亲和力结合,介导病毒包膜与细胞膜的融合,从而完成入侵过程。因此,阻断RBD与ACE2的相互作用是中和抗体发挥保护作用的主要机制,也是疫苗设计和抗体检测试剂开发的逻辑起点。
RBD作为抗原靶点具有多重优势:首先,它包含多个主要的病毒中和表位,免疫后产生的抗体能有效阻断病毒结合;其次,相较于全长S蛋白,RBD结构相对保守且分子量较小,易于通过原核或真核系统重组表达,降低生产成本。然而,随着Omicron等变异株的持续出现,RBD上的K417、L452、E484、N501等位点发生高频突变,导致病毒免疫逃逸能力增强,原有疫苗和检测试剂的效力下降。
Q2:大肠杆菌表达系统制备RBD蛋白有哪些优势与挑战?
A:大肠杆菌表达系统是重组蛋白生产中最常用、成本最低的原核表达系统。其优势包括:遗传背景清晰、操作简便、培养周期短、易于放大生产,且目标蛋白表达量高,适合工业化制备。对于RBD这类非糖基化或糖基化修饰不关键的功能域,大肠杆菌系统能够在较短时间内获得毫克至克级重组蛋白,大幅降低研发成本。
然而,大肠杆菌表达系统也面临显著挑战:由于缺乏真核翻译后修饰机制,复杂蛋白容易形成包涵体(无活性的不溶聚集体),且革兰氏阴性菌细胞壁中的内毒素(脂多糖,LPS)会强烈激活免疫细胞,影响体内外实验结果。对于RBD蛋白,如何从包涵体中复性获得正确构象的活性蛋白,并将内毒素降至安全水平,是技术难点。
Q3:E484位点突变为何能显著提升RBD蛋白的免疫原性?其机制是什么?
A:E484位点位于RBD与ACE2结合界面的边缘,是一个高度暴露的柔性环区,在多个变异株(Beta、Gamma、Kappa、Omicron)中频繁出现E484K、E484Q、E484A等突变。该位点的氨基酸改变之所以能显著提升RBD的免疫原性,主要涉及抗原结构、表位暴露和免疫识别三个层面的机制。
首先,从结构上看,野生型RBD中E484为带负电的谷氨酸,突变为带正电的赖氨酸(E484K)或中性谷氨酰胺(E484Q)会改变局部电荷分布和侧链构象,可能诱导RBD整体折叠模式的微调。这种构象变化可使得原本隐藏或半隐藏的免疫优势表位(如中和抗体结合表位)更加暴露于蛋白表面,从而增强B细胞受体的识别效率。
其次,E484突变可以改变RBD的聚集状态或降解动力学。本研究中,E484K和E484Q突变体在小鼠体内诱导的抗体滴度分别达到野生型的4.7倍和4.8倍,而K417T或N501Y单点突变则无显著提升,说明E484位点具有独特的“免疫原性增强开关”功能。这可能是因为E484突变增加了蛋白被抗原呈递细胞摄取、加工和呈递的效率,进而更有效地激活Tfh细胞和生发中心反应。
最后,研究还发现E484K突变体作为包被抗原时,能够高效识别所有免疫组血清抗体(包括野生型和多种突变体诱导的抗体),表现出优异的通用性。这说明E484突变不仅提升了自身的免疫原性,还可能诱导产生针对RBD保守区域的广谱抗体,其深层机制值得进一步通过晶体结构和B细胞表位作图研究。
Q4:RBD蛋白与ACE2的结合亲和力是否决定其免疫原性?
A:传统观念常认为,病毒抗原与宿主受体的结合亲和力越高,可能意味着该抗原表位更“关键”,从而诱导更强的免疫反应。然而,本研究通过系统比较不同RBD突变体的ACE2结合能力与体内免疫原性,认为RBD与ACE2的结合亲和力并不直接决定其免疫原性。
相反,E484K和E484Q突变对ACE2亲和力的影响相对较小(甚至在某些背景下略微降低结合力),却能大幅度提升免疫原性。这表明免疫原性主要由抗原的构象稳定性、表位暴露程度、蛋白酶体降解模式以及与先天免疫受体的互作决定,而非单纯由其受体结合能力决定。
Q5:内毒素水平对重组蛋白的体内外实验有何影响?
A:内毒素,又称脂多糖(LPS),是革兰氏阴性菌(如大肠杆菌)细胞壁的主要成分。在重组蛋白制备过程中,即使经过纯化,残留的内毒素也会对后续实验产生严重干扰。
在体外,内毒素可激活巨噬细胞、树突状细胞等,诱导TNF-α、IL-1β、IL-6等炎症因子的释放,造成Western blot背景增高、细胞活力下降(即“细胞死亡”的常见原因)。
在体内,内毒素直接激发先天免疫反应,导致小鼠发热、炎症甚至死亡,同时非特异性地增强或扭曲适应性免疫应答,使疫苗免疫原性评价结果不可靠。因此,用于动物免疫的重组蛋白,内毒素通常要求低于5 EU/mL。
- 联系我们
- 电话:400-621-6806
- 手机:13116038708(微信)
- 邮箱:info@kmdbioscience.com
- 地址:天津市武清区国际企业社区A5号楼
-

销售微信
-

官方公众号
-
 400-621-6806
400-621-6806 -
 3599163699
3599163699 -
 周一至周五 9:00-18:00
周一至周五 9:00-18:00 -
-
 0
0
-
