

设计“完美抗原”:mRNA免疫原工程的核心策略
- 2026-07-14
- 11
上一篇讲到了mRNA免疫的底层逻辑,设计mRNA编码的核心目标是以此精准引导免疫系统,诱导产生目标抗体。抗原设计的最终目标是让重组蛋白处于合适的位置,能够有符合的表位构象呈现给免疫系统,以足够的产量构建出合适的结构状态。
一、提升转录翻译的效率——产量
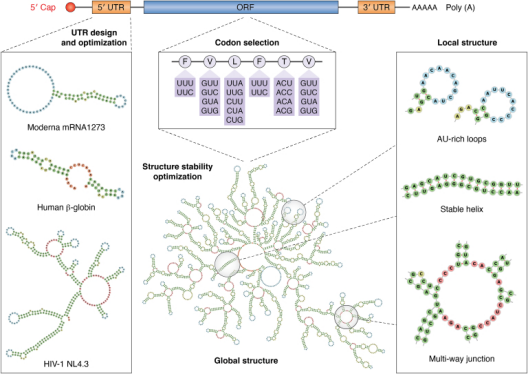
1、5′UTR:
设计5′UTR最直接的方法是从已知的现有人类mRNA中选择5′UTR序列,并在目标细胞类型中具有较高的表达效率。即便如此天然衍生的5′UTR可以包含多种调节元器件,也可能会对特定条件下的核糖体负荷产生阻碍作用[1]。所以在设计mRNA的5′UTR时应遵循在减少抑制性结构(如稳定发卡、uORF)的同时优化翻译起始效率的策略。
设计工具:这几年,越来越多的预测模型涌现出来,专门用来模拟5′UTR对蛋白质表达的影响,并进一步优化和设计5′UTR序列。现在主流的方法包括随机森林(Random Forests,RF),卷积神经网络(Convolutional Neural Networks,CNN),和对抗生成网络(Generative Adversarial Networks,GAN)等方法[1]。与此同时,研究者们还建立了不少关于5′UTR的专门数据库,直接为这些模型的训练和评估提供了数据支持。
2、3′UTR和Poly(A)尾:
虽然3′UTR虽然调控mRNA稳定性和翻译效率上不及5′UTR关键,但其蕴含的调控元件仍然十分丰富。例如富含AU的元件(AU-rich Element,ARE)和微小RNA(microRNA,miRNA)结合位点。
Poly(A)尾调控mRNA的定位、稳定性和翻译中发挥关键调控作用。其长度是影响内源性mRNA和外源性递送mRNA的稳定性及翻译效率的重要参数[2]。随着TAILseq、mTAILseq、PALseq、FLAMseq、PATseq及纳米孔测序等高通量测序技术的发展,目前研究者能够对Poly(A)的长度和序列组成进行深入分析[3]。
3、密码子优化:
密码子优化是将理想密码子为每个氨基酸选择的过程。同义密码子会对mRNA的翻译效率和稳定性产生影响[4]。一种通常采用高丰度的转运RNA(Transfer RNA,tRNA)对应的偏好密码子替代稀有密码子来实现翻译速率的提升,此种密码子使用偏差可以通过密码子适应指数(CAI)来量化[1]。
密码子优化通常要求CAI值优化至宿主偏好范围内,避免极端偏置,另外在设计过程中也需避免重复序列导致的聚集引发核糖体移码。
二、抗原的亚细胞定位设计——定位
抗原最终处在的位置,直接决定了他被免疫细胞“看见”的方式,从而塑造其体液免疫和细胞免疫的平衡。
1、分泌型:
分泌型抗原设计是将一段可切割的信号肽融合在抗原的N端,经高尔基体成熟后,经分泌囊泡释放到细胞外,将新生肽链引导到内质网(ER)[5]。促使目标抗原直接释放于培养上清中,此种策略不仅简化了纯化流程,并且主要诱导偏向Th2型的强效体液免疫应答。
2、膜结合型:
通过对跨膜区(Transmembrane Domain,TMD)的保留和移植,将抗原锚定于细胞膜表面。常见的膜结合型抗原有设计骨架有:Ⅰ型跨膜蛋白的N端位于胞外、C端位于胞内[6]。这种方法更好地模拟了天然病毒的状态,有利于诱导细胞免疫应答。
3、胞质型:
通过移除信号肽并借助N端碱性残基的胞质锚定作用,使抗原在细胞质中有效滞留。这种策略适用于胞内抗原疫苗,其通过MHCⅠ类分子呈递,强力激活细胞活性。
三、抗原高级结构合理设计
以上两个模块完成之后,我们获得了在正确位置高表达的蛋白,然而,高表达并不等同于高功能——蛋白是否正确折叠、关键表位是否充分暴露,仍需进一步验证。更关键的是,该抗原能否在动物免疫过程中有效激发目标抗体应答,直接决定了后续抗体发现工作的成败。无论最终目标是获得多克隆抗体用于诊断,还是通过噬菌体展示或杂交瘤技术进行单克隆抗体的制备,抗原的构象真实性、表位可及性与多聚化程度都是核心前提。因此,抗原高级结构的合理设计必须服务于特定的免疫策略和抗体筛选路线。
1、构象设计:
利用PDB或AlphaFold结构,通过引入二硫键或疏水核心优化提升热稳定性,以保证该蛋白在37℃环境下维持三聚体形态,因为只有保持天然三聚体构象,才能保证其正确的抗原表位暴露,从而诱导有效的功能性抗体。
2、表位聚焦:
可以通过丙氨酸扫描定位免疫优势的表位,并引入点突变破坏其与抗体的结合力,从而清除免疫显性但非保护性的表位。也可以对此进行糖基化屏蔽,即在非中和表位邻近区域插入N-X-S/T基序,利用聚糖的空间位阻掩盖区域,迫使其免疫应答转向保守的中和表位。
3、展现形式:
①铁蛋白纳米笼:将抗原单体融合至铁蛋白N端,将关键表位置于纳米颗粒表面,与B细胞受体进行强烈交联。
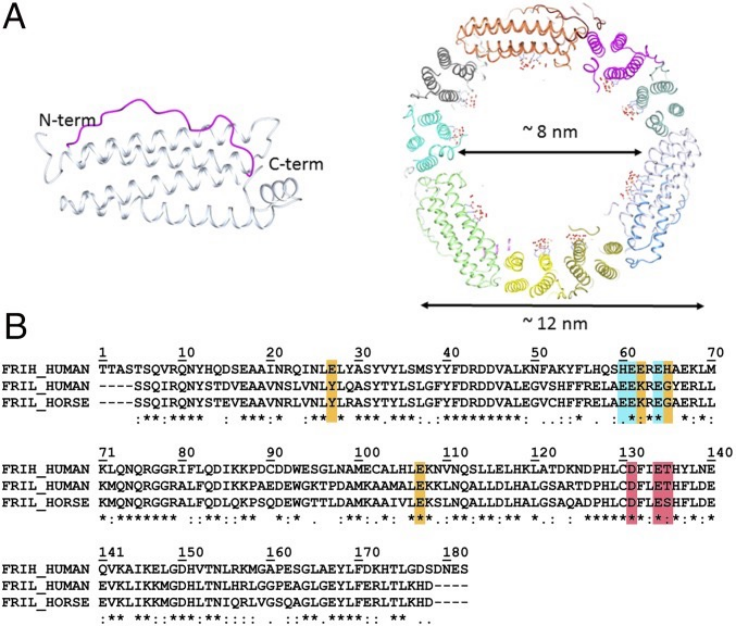
②类病毒颗粒:利用外壳蛋白作为骨架,将目标表位嵌合在颗粒表面暴露的环区,可以借助颗粒尺寸优先被淋巴结捕获。
四、验证效果的工具和方法
1、表达与定位的快速初筛:
① 蛋白质免疫印迹(Western Blot,WB):检测全细胞裂解液和上清液,可快速确认分子量是否正确,是否完整表达,有无降解情况。
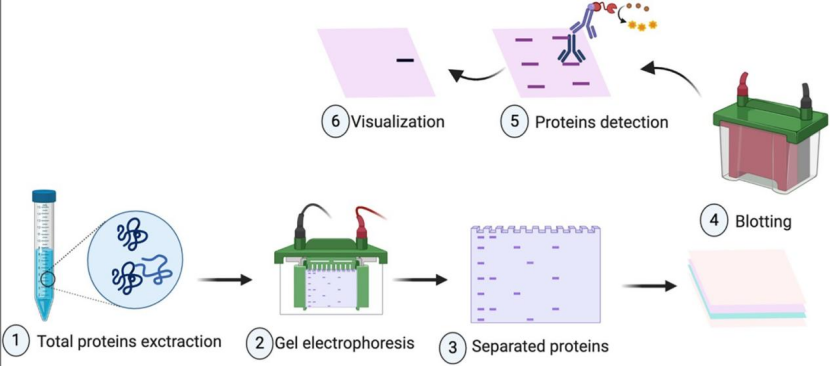
②免疫荧光:用特异性标签与细胞器标记物共染,精准判定蛋白定位。
2、构象与表位可及性验证:
①酶联免疫吸附试验(Enzyme-Linked Immunosorbent Assay,ELISA):使用识别线性表位的抗体作为对照与识别天然构象表位的抗体进行实验验证。若构象抗体结合信号/线性抗体结合信号 > 阈值,则说明构象正确。
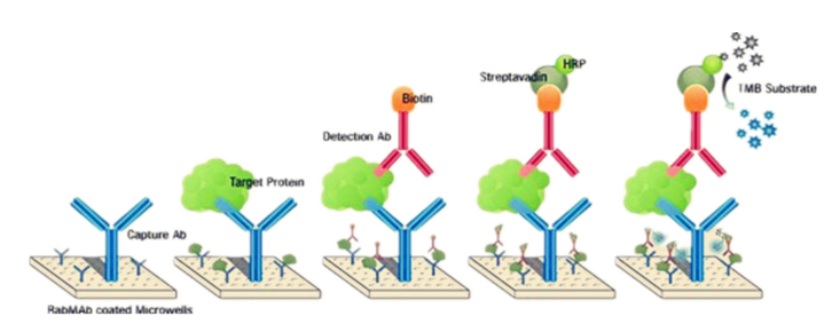
图4:ELISA方法测定原理图
②流式细胞荧光分选技术(Fluorescence Activated Cell Sorter,FACS):为评估膜结合型抗原的天然构象表位暴露情况,利用流式细胞术检测中和抗体的结合能力。若实验结果显示,荧光强度随抗体浓度增加而呈现梯度饱和,便可证实靶标表位在活细胞表面充分暴露且可及。
在抗原生产端,卡梅德生物提供大肠杆菌、酵母、昆虫细胞及哺乳动物细胞等多种表达系统的重组蛋白表达定制服务,涵盖各类受体、细胞因子、病毒抗原等;在构象验证端,可提供从基因序列分析到抗体纯化的一站式抗体制备服务,包括人源化抗体、纳米抗体等服务。
从抗原设计到蛋白生产,从抗体开发到细胞系构建——卡梅德生物用一站式的技术平台,助您把“完美抗原”的设计蓝图,变成实验室里可触可用的科研利器。
本文系统梳理了mRNA免疫原工程的全链条策略:从5′UTR、密码子优化到Poly(A)尾设计,提升抗原产量;通过分泌型、膜结合型或胞质型定位,调控免疫应答方向;进而利用构象锁定、表位聚焦及纳米颗粒多聚化展示,确保正确表位呈递。配套ELISA、FACS等验证方法,为高效抗体诱导提供可落地的设计框架,最终指向优化生发中心应答、实现广谱持久免疫的目标。
当我们通过上述策略设计出能展示“完美状态”的mRNA之后,最终目标为建立高效、持久、广谱的抗体应答,这个目标如何实现?体内的生发中心反应如何被优化?mRNA是否能直接用于编码治疗性抗体?最后一篇将完整解答。
参考文献
[1]Jin L, Zhou Y, Zhang S, Chen SJ. mRNA vaccine sequence and structure design and optimization: Advances and challenges. J Biol Chem. 2025;301(1):108015. doi:10.1016/j.jbc.2024.108015
[2]Spiewla T, Czubak K, Pilch Z, et al. PolyA tail segmentation improves the stability of the template DNA and increases the translatability of in vitro transcribed mRNA. Nucleic Acids Res. 2026;54(2):gkaf1412. doi:10.1093/nar/gkaf1412
[3]Passmore LA, Coller J. Roles of mRNA poly(A) tails in regulation of eukaryotic gene expression. Nat Rev Mol Cell Biol. 2022;23(2):93-106. doi:10.1038/s41580-021-00417-y
[4]Li Y, Wang F, Yang J, Han Z, Chen L, Jiang W, Zhou H, Li T, Tang Z, Deng J, He X, Zha G, Hu Z, Hu Y, Wu L, Zhan C, Sun C, He Y, Xie Z. Deep generative optimization of mRNA codon sequences for enhanced mRNA translation and therapeutic efficacy. Nat Commun. 2025 Nov 12;16(1):9957. doi: 10.1038/s41467-025-64894-x. PMID: 41224770; PMCID: PMC12612108.
[5]Sato S, Minagawa N, Hirata Y, et al. Evaluating Signal Peptide Efficiency for Extracellular Protein Secretion for mRNA Vaccine Design. Biol Pharm Bull. 2025;48(5):706-712. doi:10.1248/bpb.b25-00155
[6]Sueda S, Tsuruga R, Hirakawa T, Fujii S. Cell surface display of a protein based on a tail-anchored membrane protein. Biochem Biophys Res Commun. 2025;761:151738. doi:10.1016/j.bbrc.2025.151738
[7]Pozzi C, Ciambellotti S, Bernacchioni C, Di Pisa F, Mangani S, Turano P. Chemistry at the protein-mineral interface in L-ferritin assists the assembly of a functional (μ3-oxo)Tris[(μ2-peroxo)] triiron(III) cluster. Proc Natl Acad Sci U S A. 2017 Mar 7;114(10):2580-2585. doi: 10.1073/pnas.1614302114. Epub 2017 Feb 15. PMID: 28202724; PMCID: PMC5347543.
FAQs
1.文章中提到5′UTR设计时,天然衍生的UTR可能含有阻碍核糖体负荷的元件。具体来说,哪些类型的调控元件是需要被剔除的“多余”元件?
主要需要关注两类:一是上游开放阅读框 ,它们会竞争性消耗扫描中的核糖体,降低主开放阅读框的翻译起始效率;二是二级结构过于稳定的区域,尤其是靠近5′UTR附近的颈环结构,会物理性地阻碍核糖体43S前起始复合物的扫描。此外,某些细胞类型特异性的抑制性调控基序也需要根据目标细胞系进行针对性剔除。
2.关于Poly(A)尾长度,文章提到“长度是重要参数”。在实际mRNA设计中,是否存在一个最优的poly(A)长度区间?
目前主流观点认为,100~120个腺苷酸(A) 是较为理想的长度区间。过短的poly(A)尾(<30 nt)无法有效结合PABP(poly(A)结合蛋白),导致mRNA迅速脱腺苷酸化并降解;而过长的poly(A)尾(>200 nt)反而可能引发翻译抑制,且会增加质粒构建和IVT(体外转录)的难度。不过,最优长度也可能因细胞类型和递送方式而略有差异,建议在设计时参考同类抗原在目标体系中的经验数据。
3.密码子优化中,CAI值不低于0.85是一个常见标准。如果某个抗原序列的CAI值已经很高,是否还有必要关注mRNA二级结构?
非常有必要。CAI衡量的是密码子使用频率与宿主tRNA丰度的匹配程度,但它并不直接反映mRNA的可翻译性 。即使CAI很高,如果mRNA在起始密码子AUG周围形成了高度稳定的发夹结构,核糖体仍然无法有效结合并启动扫描,翻译效率会大打折扣。因此,CAI与mRNA二级结构自由能必须协同优化,两者不可偏废。
4.文章中“构象设计”部分提到通过引入二硫键使Tm值提升5~10℃。为什么Tm值提升这个幅度具有生物学意义?
人体核心温度约为37℃,而重组抗原在体内面临的温度环境可能更高。Tm值代表蛋白从天然折叠态到去折叠态的转变中点温度,将Tm值从40℃提升至45℃以上,意味着蛋白在37℃下的去折叠比例呈指数级下降。这使得抗原在体内数小时至数天的暴露窗口期内,能够持续以正确构象呈递给免疫细胞,而非迅速变为无功能的聚集态。
5.铁蛋白纳米笼和类病毒颗粒(VLP)都用于多聚化展示,两者在设计思路上有何本质区别?
两者的核心区别在于展示逻辑。铁蛋白纳米笼属于“人工合成支架”即抗原单体通过基因融合或化学偶联的方式连接在铁蛋白亚基的N端或C端,组装后抗原被“悬挂”在24聚体纳米颗粒的表面。这种方式的优势是结构规整、尺寸均一,且不依赖病毒来源的组分。VLP则是“天然来源骨架”即利用病毒的结构蛋白自组装形成颗粒,将目标表位通过基因工程插入到外壳蛋白表面暴露的环区。VLP的优势在于保留了病毒颗粒的天然免疫刺激性,但骨架本身可能引发抗载体免疫反应,多次免疫时需考虑载体抑制效应。
- 联系我们
- 电话:400-621-6806
- 手机:13116038708(微信)
- 邮箱:info@kmdbioscience.com
- 地址:天津市武清区国际企业社区A5号楼
-

销售微信
-

官方公众号
-
 400-621-6806
400-621-6806 -
 3599163699
3599163699 -
 周一至周五 9:00-18:00
周一至周五 9:00-18:00 -
-
 0
0
-
